I took a course titled, "Mathematical Ecology" in undergrad at the University of Texas. The course was taught by
Dr. Eric Pianka and it was the worst classes I ever took. Well, no, not really. It was a distressing class, though. I studied like mad and got Ds on every mid-term. I didn't realize until the night before the final how I was approaching the class wrong. Most points in each midterm were assigned on the basis of questions about a single mathematical model described in the course section (and not all the other little info-bits that I had been focusing on). I stayed awake all night, studying only the mathematical models discussed all semester. I ended up getting a B as my final grade for the semester.
I never did pick up the graded exam, but my course grade told I ended up blowing the exam out of the water. Now, this alone might not have been enough to get the grade I got. I suspect the professor acknowledged my --eventual-- mastery of the semester's most important material by making my final exam score have a bigger impact in the overall grade than it was originally designed to be. Thanks very much for that, Dr. Pianka.
 |
Figure 1a. Positive
interactions. |
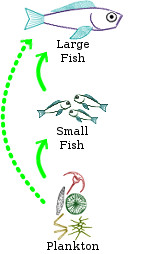
One particular lesson started with an observation of life out in the open ocean, well away from shore or reef systems at least. Out in the open ocean, there is a large amount of biomass present in tiny plankton and in large fish, but relatively little in small fish. The plankton feeds the small fish and then the small fish feed the large fish. Another way to say this is that plankton biomass has a positive influence on small fish biomass and small fish biomass has a positive influence on large fish biomass. We can represent each positive interaction with a pointed arrow. One positive influence and a second positive influence together forms an overall positive influence (Figure 1a). This can be translated into an equation as: (I
1 * I
2) = I
f.
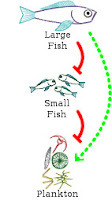 |
Figure 1b. Negative
interactions. |
We can think about these steps in the opposite direction. Large fish biomass has a negative influence on the amount of small fish biomass, which has a negative influence on the amount of plankton biomass. We can represent each negative interaction with a flat-arrow, going the opposite direction of the earlier pointed arrows. One negative influence and a second negative influence together also forms an overall positive influence (Figure 1b). This can be translated into an equation as: (-I
1 * -I
2) = I
f.
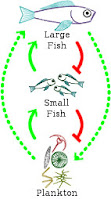 |
Figure 1c. Fishy
feedback. |
Plankton biomass has a positive influence on large fish biomass and large fish biomass has a positive influence on plankton biomass. All together (Figure 1c), we have a positive feedback loop that explains why there is more biomass in plankton and large fish than there is in small fish. On average, the ecosystem only produces enough small fish to be consumed by the large fish. Any more small fish would result in the growth of the population of large fish. Any less small fish would result in a reduction in the population of large fish.
This sort of analysis won't let you know what levels the elements of the system being modeled will stabilize at, or if it will even stabilize at all, but it can quickly give you an overview of the likely overall behavior of the system.
 |
Figure 2a. Otters in
the Kelp forest. |
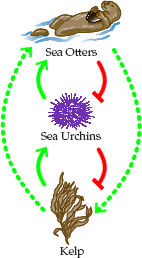
Another scenario with the same dynamics is the relationship between Kelp, Sea Urchins, and Sea Otters (Figure 2a). This relationship is discussed in some detail in a
recent episode of Science Friday. Otters help maintain the kelp forest by limiting the numbers of the major herbivore (urchins) which would otherwise
obliterate the kelp.
 |
Figure 2b. Orca in
the Kelp forest. |
We can add Orca, a major predator of sea otters, into the model (Figure 2b). Because Sea Otters aren't the dominant food source for Orca, I haven't added a positive arrow between Sea Otters and Orca. The model allows us to see that Orca have a negative influence on Kelp forests. Humans hunting otters for their fur would have a greater impact than the Orca because the humans are looking specifically for the Sea Otter (no matter how scarce), while the Orca will
switch to other prey if Sea Otters become scarce.
A more complicated system also discussed in
the episode of Science Friday describes how the introduction of a vaccine against the Rinderpest virus could have an impact on the population of Giraffes on the Serengeti. Rinderpest is a virus of cattle which kills Wildebeest. Wildebeest eat large amounts of grass, so when Wildebeest are killed off, more grass will grow. More grass means the ecosystem is more susceptible to fires, which kill Acacia seedlings. Since adult Acacia trees feed Giraffes, more fires means less food for Giraffes. Altogether, the model presented in Figure 3 shows that the introduction of the Rinderpest vaccine would result in an increased population of Giraffes. Unfortunately, this analysis wasn't done in advance of vaccine introduction. Instead it was developed in an attempt to understand why the population of Giraffes increased after the introduction.
 |
| Figure 3. Impact of Rinderpest vaccine on Serengeti Giraffes. |
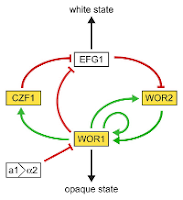 |
Figure 4a. Opaque-white
switching in C. albicans. |
The same logic can be used to analyze networks of interactions outside ecological modeling. Figure 4a describes the genetic regulatory network responsible for white-opaque switching in
Candida albicans, a sometimes pathogenic yeast commensal of the human gut. Opaque-white switching refers to a transition of the yeast between a reproductive and vegetative developmental state. The opaque/white labels refer to how colonies of the cells in each developmental state appear when growing on media in a petri dish. My Figure 4a is updated a figure from
the original paper describing this interaction network so it has colors consistent with my other figures here. The relationships illustrated in the figure had been constructed by involved experiments examining interactions between each of the genes involved in the process.
The diagram was presented by a speaker at a yeast research conference I attended. The speaker then went through an involved discussion of all the experiments they did to explain why the network resulted in the final observed behavior of switching between the white and opaque developmental states. As the presentation went on and on, I found myself getting more and more irritated. I was expecting a couple slides describing the interactions, then a continuation on to the real meat of the presentation. It was quickly clear to me why this network would result in the observed switching behavior because I was analyzing it using the techniques discussed earlier in this posting. The final conclusion of the presentation was the result I understood at the beginning of the presentation. I felt very frustrated.
After the presentation, I tried to convey my frustration to my graduate advisor. Unfortunately, I didn't know how to explain how I was processing the interaction network that had been shown. It turns out that most biologists don't have the mathematical background that I had taken for granted in my own research.
 |
| Figure 4b. Simplification step 1. |
I knew how the overall network behaved because I was able to quickly simplify the network. In the left half of the network (in Figure 4a), the positive interaction between WOR1 and CZF1 and the negative interaction between CZF1 and EFG1 together have an overall negative interaction. There is already a negative interaction between WOR1 and EFG1, so this side chain would act to amplify the effect. For our purposes, this means we can simplify the network by removing this side chain (Figure 4b).
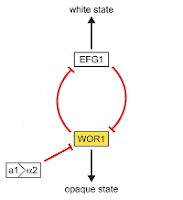 |
| Figure 4c. Simplification step 2. |
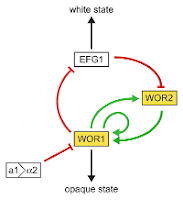
The interactions between WOR1 and WOR2 represents a feedback loop which stabilizes them in an active state. The negative interaction between EFG1 and WOR2, paired with the positive feedback loop, means that the right half of the figure has an overall negative influence between EFG1 and WOR1. Again, we can simplify the network by drawing a single negative interaction on the right (Figure 4c).
Two elements in the network with negative interactions between them represent a bistable switch. If either WOR1 or EFG1 gets the upper hand, it then suppresses the other. Only one will ever be active at one time. The final element of the network which remains is the influence of the a1/α2 ratio, which acts as an outside control element on the switch. There are other elements impacting the activity of EFG1 which aren't described in this network, but they will also act as external control elements. Together, these outside influences determine the state of the switch and when it can change from one state to the other.
The relationship between WOR1 and EFG1 was very quickly apparent to me at the start of the presentation, but then the presentation kept going on and on in some detail trying to explain the overall behavior. A the time of the conference, my frustration then was in why the presenter wasted so much of our time, as well as in my inability to explain this concept to my graduate advisor.
Now, however, my frustration with this memory has more to do with my irritation at researchers not collaborating with mathematicians (or at least mathematically-inclined researchers) during their work. Having some basic level of literacy at math, computer programming, statistics, or any other specialized field can help you in many ways. You don't need to be an expert in these subjects because even a limited grasp of them will help you to know when it is a good idea to call in someone with the specialized knowledge that you lack.
Dr. Pianka's course is the one I have most
often thought of over the years. Specific lessons I learned in his course routinely come
to mind and help guide how I interpret material seen in my scientific career so far. I expect the course will continue to inform my future biology research endeavors. Thank you again, Dr. Pianka.
References
 I just returned from a week of traveling in Nevada and California. We visited with my wife's grandmother for a few days, then headed out on the road. We stopped by the Luther Burbank house and hiked through several botanical gardens. The inevitable consequence of this trip is that I took many photos (of plants, animals, and geology) that will be appearing in posts over the coming months.
I just returned from a week of traveling in Nevada and California. We visited with my wife's grandmother for a few days, then headed out on the road. We stopped by the Luther Burbank house and hiked through several botanical gardens. The inevitable consequence of this trip is that I took many photos (of plants, animals, and geology) that will be appearing in posts over the coming months.